Bacterial Defence Mechanisms and Bacteriophage Counter-defence Mechanisms Nature Review Microbiology
![]() Open access peer-reviewed chapter
Open access peer-reviewed chapter
The War betwixt Leaner and Bacteriophages
Submitted: July 15th, 2018 Reviewed: June 4th, 2019 Published: October 31st, 2019
DOI: 10.5772/intechopen.87247

IntechOpen Downloads
1,505
Total Chapter Downloads on intechopen.com
![]()
Altmetric score
Overall attention for this chapters
Abstract
The rapid emergence and broadcasting of multidrug-resistant (MDR) bacteria represents a worldwide crunch apropos that humankind is re-entering the 'pre-antibiotics' era. Earlier the discovery of antibiotics, bacteriophage therapy was widely enforced to combat bacterial infections. However, the discovery of penicillin in 1940 and other novel antibiotics replaced phage therapy, and they are being used as the kickoff line of defence confronting pathogenic bacterial infections. Factors such as selective force per unit area resulted in bacteria condign insensitive to one or multiple antibiotics, often leading to limited handling options. This prompted a renewal of involvement to the phage therapy that remains dubious due to its disadvantages such every bit host specificity and the development of bacterial resistance against phages. Evolution of bacterial genomes immune bacteria to acquire vast mechanisms interfering with phage infection such as inhibition of phage adsorption, prevention of phage entry, superinfection exclusion, restriction-modification and abortive infection. Interestingly, phages accept developed diverse counterstrategies to circumvent bacterial anti-phage mechanisms including excavation for receptors, adapting to new receptors and masking and modifying restriction sites. Understanding the complex dynamics of bacteria-phage interaction is a preliminary step towards designing synthetic phages that can overcome limitations of phage therapy and potentially lead to defeating MDR bacteria.
Keywords
- bacteria-phage arms race
- CRISPR system
- anti-CRISPR system
- superinfection exclusion (Sie)
- restriction-modification
- abortive infection (Abi)
*Address all correspondence to: m.mohammed@westminster.ac.uk
1. Introduction
Antimicrobial resistance is a global public health crisis. According to Public Health England [one], each year approximately 25,000 people die beyond Europe due to hospital-acquired infections caused past antibiotic-resistant and MDR bacteria such every bit
Félix d'Herelle, known as the father of bacteriophage (or phage) therapy [three], brought an evolutionary discovery of phages equally therapeutics for various infections and conditions. Phage therapy was widely enforced in the 1920s and 1930s to gainsay the bacterial infections. However, in the 1940s, the newly discovered antibiotics replaced the phage therapy (except Russia, Georgia and Poland) [four].
The emergence of MDR leaner prompted a renewal of the involvement to the phage therapy as an alternative handling to overcome a wide spectrum of resistant bacterial infections. Phage therapy and phage cocktails that contain a mixture of different leaner-specific phages, fatigued interest within molecular biology and modern medical enquiry as potential antimicrobials that could tackle the crunch of antimicrobial resistance. Nonetheless, the phage therapy remains controversial due to its disadvantages such as bacteriophage resistance: bacteria-phage evolutionary arms race that could put a burden on a long-time awarding of phage therapy equally an anti-infectious amanuensis [5].
Phage therapy has many advantages, primary because phages are very specific (generally express to 1 species) and easy to obtain as they are widely distributed in locations populated by bacterial hosts including soil and seawater, and they do not have any known chemic side effects similar antimicrobials [6].
Agreement host-phage interactions and 'the war between bacteria and phages' are steps towards designing engineering science 'broad-spectrum phage' that can overcome the limitations of phage therapy and potentially overcome a broad range of resistant bacterial infections [6].
Advertisement
2. The evolutionary phage-host arms race
Phages are obligate intracellular parasites that distinctively infect bacterial cells. Although phages are very specific to their host, generally limited to ane species, they pose an enormous threat to bacteria as in some habitats they outnumber their hosts by nearly 10-fold number [7]. Phages are the near abundant, ubiquitous and diversified organisms in the biosphere [8, 9]. Phage-host interaction and fight for the survival led to the development of bacterial and viral genomes and, therefore, to the evolution of resistance mechanisms. Bacteria, continuously, evolve many molecular mechanisms, driven by gene expression to preclude phage infection. These evolving phage-resistance mechanisms in bacteria induce the parallel co-development of phage multifariousness and adaptability [10, 11]. The co-evolving genetic variations and counteradaptations, in bacteria and phages, drive the evolutionary phage-host arm race [eleven, 12].
Leigh Van Valen, an evolutionary biologist, metaphorised the co-evolutionary arm race and proposed the Red Queen hypothesis [thirteen].
'It takes all the running you lot can do, to stay in the same place' the Red Queen says to Alice in
The Red Queen hypothesis proposes that to survive, microorganisms must constantly adapt, evolve and thrive confronting ever-evolving combative microorganisms inside the same ecological niche [14].
Bacteria have developed various anti-phage mechanisms including non-adaptive defences (non-specific) and adaptive defences associated with Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) along with CRISPR-associated (Cas) proteins [7, 15, 16, 17, 18].
The non-specific adaptations (analogues to innate immunity in multicellular organisms) act every bit master mechanisms to evade viral infection, and they include mechanisms that inhibit phage adsorption and prevent nucleic acid entry, superinfection exclusion systems, restriction-modification systems and abortive infection [7, xix].
On the other manus, the adaptive resistance (analogues to the acquired immunity in multicellular organisms) serves as a second line of defense, which is very efficient and phage-specific.
Interestingly, it was observed that the bacterial anti-phage mechanisms are generally present in a genomic array, known equally 'defence islands' [20]. The 'defense force islands' are enriched in putative operons and contain numerous overrepresented genes encoding diverged variants of antiviral defence systems. Moreover, scientific evidence and characteristic operonic organisation of 'defense islands' show that many more than anti-phage mechanisms are yet to be discovered [21, 22, 23, 24].
Although leaner have developed several resistance mechanisms against phages, phages can circumvent bacterial anti-phage mechanisms on the grounds of their genomic plasticity and rapid replication rates. These counterstrategies include point mutations in specific genes and genome rearrangements that allow phages to evade bacterial antiviral systems such every bit CRISPR/Cas arrays past using anti-CRISPR proteins and abortive infection by hijacking bacterial antitoxins, equally well as escaping from adsorption inhibition and restriction-modification mechanisms [15, 16, 17, 18].
This affiliate will comment on the genetic basis of bacterial resistance to phages and unlike strategies used by phages to evade bacterial resistance mechanisms.
Advertisement
3. Preventing phage adsorption and phage's counterstrategy
Phage adsorption to host-specific receptors on the jail cell surface is the initial step of the infection and host-phage interaction. Depending on the nature of bacteria, whether it is Gram-positive or Gram-negative proteins, lipopolysaccharides, teichoic acids and other cell surface structures tin serve as irreversible phage-binding receptors [19]. These receptors might be present in the prison cell wall, bacterial capsules, slime layers, pili or flagella [25].
Leaner have acquired various barriers to inhibit phage adsorption, such every bit blocking of phage receptors, product of extracellular matrix (e.g. sheathing, slime layers) and production of competitive inhibitors [26, 27, 28, 29, 30, 31]. The diverseness of phage receptors in the host is influenced by co-evolutionary adaptations of phages to overcome these barriers [32]. This includes diversity-generating retroelements (DGRs) and phase variation mechanisms causing phenotypical differences within the bacterial colony [7, 33, 34].
Phase variation is a heritable, yet reversible process regulating factor expression in leaner; genes can switch between a functional (expression) and a not-functional state leading to phenotypical variations within the bacterial population even when strains have identical genotype. Sørensen et al. [35] investigated the underlying resistance mechanism of
DGRs are genetic elements diversifying DNA sequences and the proteins they encode ultimately mediating the development of ligand-receptor interactions. Fault-prone DGRs and random mutations in the bacterial genes encoding prison cell surface receptors lead to the alternation and change in the structural composition of the phage receptors, making them non-complementary to the phage'due south anti-receptors, known as receptor-bounden proteins (RBP) [34] (Figure ane(1)).
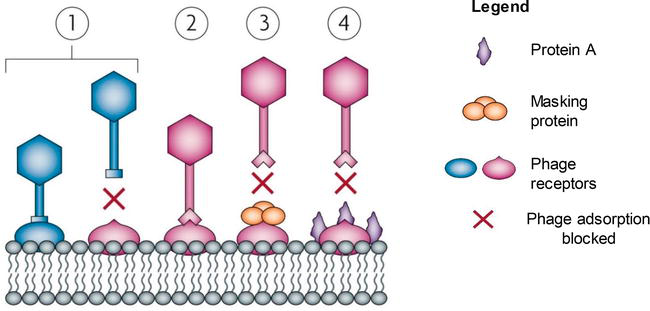
Figure i.
Bacterial defence force mechanisms preventing phage adsorption and phage'due south counteradaptations. (ane) Phage adsorption to a host-specific receptor site on a host cell surface. Bacterium evolves phage resistance past the modification of these cell surface receptors; phage is incapable of binding to the altered receptor. (ii) Phage's adaptation to these modifications through mutations in receptor-binding protein cistron that leads to the co-development of bacterial genetic variation. Leaner are as well capable of producing proteins that mask the phage recognition site receptors (3 and 4), thus making the receptor inaccessible for phage adsorption [
All the same, phage'due south replication is exceedingly fault-prone, therefore causing many random mutations in the genes encoding the RBP or tail fibres. Phages also possess DGRs that mediate phage'southward tropism by accelerating the variability in the receptor-coding genes through reverse transcription process [37]. The changes in the nucleotide sequence in the RBP-coding factor may ultimately atomic number 82 to the adaptation to the modified receptor (Figure 1(2)), thus the power to adsorb and infect the bacterial prison cell.
Unsurprisingly, leaner as well exhibit different strategies to block their receptors [28, 29, 30, 31].
Figure 1(4) demonstrates the findings from studies conducted on
Receptors located on bacterial cell surface serve a vital office in bacterial metabolism; they may function as membrane porins, adhesions or chemical receptors [19]. Therefore, mutation or consummate loss of the receptor might exist lethal for leaner. To inhibit phage adsorption, bacteria tin can produce surface molecules, such as exopolysaccharides.
Exopolysaccharides are extracellular polysaccharides interim as a concrete barrier, composing slime or capsules surrounding bacterial cells that lead to inaccessible host receptors for efficient phage adsorption [39] (Figure 2). Studies conducted by Looijesteijn et al. [40] shown that exopolysaccharides
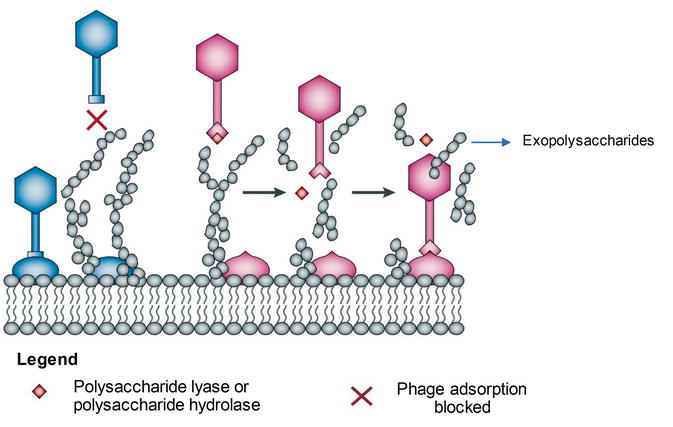
Figure 2.
Bacterial strategies to inhibit phage adsorption and phage strategies to admission host receptors. Some bacteria are capable of the production of exopolysaccharides, which human activity equally an outer shield, protecting a cell from the phage infection [
Notwithstanding, some phages evolved mechanisms allowing them to recognise these extracellular matrixes and dethrone them by utilising hydrolases and lyases (Figure ii) [15, 16, 17, 18]. The polysaccharide-degrading enzymes allow phages to gain access to the receptor that may lead to the viral propagation. They are commonly present leap to the RBPs or exist as free soluble enzymes from previously lysed bacterial cells [41].
Advertizing
4. Preventing phage DNA entry and phage's counteradaptations
If phage bypasses primary antiviral strategies, it is now able to initiate infection by adsorption to a specific receptor site on a host cell surface through phage RBP [42, 43]. Upon interaction with the cell receptors, the phage injects its genetic textile (unmarried or double-stranded DNA or RNA) into the cytoplasm of the host. Depending on the nature of the phage and growth atmospheric condition of the host jail cell, it follows one of the two life cycles: lytic or lysogenic (Figure 3).

Effigy 3.
Lytic and lysogenic life cycles of a temperate coliphage λ that infects
In the lytic cycle, virulent phages degrade host'south genome leading to the biosynthesis of viral proteins and nucleic acids for the associates of phage progeny. Somewhen, the bacterial cell lysis, releasing a multitude of newly assembled phages, is set to infect a new host prison cell [46].
In contrast, temperate phages might enter the lytic or lysogenic wheel, if the host cell exists in agin ecology weather that could potentially limit the number of produced progeny (Effigy iii demonstrates typical lifecycle of temperate phage using coliphage λ equally an case) [44, 45]. In the lysogenic phase, repressed phage genome integrates into the bacterial chromosome equally a prophage. This procedure causes the proliferation of prophage during replication and binary fission of bacterial DNA.
Prophage just expresses a repressor protein-coding gene. The repressor poly peptide binds to the operator sites of the other genes and ultimately inhibits synthesis of phage enzymes and proteins required for the lytic cycle.
When the synthesis of the repressor protein stops or if it becomes inactivated, a prophage may excise from the bacterial chromosome, initiating a lytic cycle (consecration) which leads to the multiplication and release of virulent phages and lysis of a host cell [44, 45].
If the phage remains in the nearly dormant country (prophage), the lysogenic bacterium is allowed to subsequent infection by other phages that are the same or closely analogous to the integrated prophage by means of Superinfection exclusion (Sie) systems [47].
Sie systems are membrane-associated proteins, mostly, phage or prophage encoded, that preclude phage genome entry into a host cell [47]. Figure 4 shows the office of Sie system (proteins Imm and Sp) in blocking phage T4 DNA entry into Gram-negative

Figure four.
Superinfection exclusion systems preventing phage Dna entry in Gram-negative
Advertizing
five. Host strategies to cleave invading genomes and evolutionary tactics employed by phages to featherbed these antiviral mechanisms
The development of bacterial genomes allowed bacteria to acquire vast mechanisms interfering with every stride of phage infection. In a case where a phage succeeded to inject its viral nucleic acid into a host cell, bacteria possess a multifariousness of nucleic acrid degrading systems such every bit restriction-modification (R-K) systems and CRISPR/Cas that protect bacteria from the phage invasion.
5.ane Restriction-modification systems
It has been reported that R-M systems tin can significantly contribute to bacterial resistance to phages [49].
R-M systems incorporate activities of methyltransferases (MTases) that catalyse the transfer of a methyl grouping to DNA to protect cocky-genome from a restriction endonuclease (REase) cleavage and REases, which recognise and cut strange unmethylated double-stranded DNA at specific recognition sites, commonly palindromic. To protect self-DNA from the degradation, methylases tag sequences recognised by the endonucleases with the methyl groups, whereas unmethylated phage (nonself) Deoxyribonucleic acid is cleaved and degraded (Figure 5) [26, 27, 50, 51, 52].

Figure 5.
General representation of the bacterial restriction-modification (R-M) systems providing a defence confronting invading phage genomes. R-M systems consist of two contrasting enzymatic activities: a brake endonuclease (REase) and a methyltransferase. REase recognises and cuts nonself unmethylated double-stranded DNA at specific recognition sites, whereas MTase adds methyl groups to the same genomic recognition sites on the bacterial DNA to protect cocky-genome from REase cleavage [
R-1000 systems are various and ubiquitous amid bacteria. There are iv known types of R-Chiliad within bacterial genomes (Figure 6). Their classification is mainly based on R-M system subunit composition, sequence recognition, cleavage position, cofactor requirements and substrate specificity [26, 27, 50, 51].

Effigy 6.
Four distinct types of brake-modification (R-M) systems. (a) Type I R-M system is composed of 3 subunits forming a complex: hsdR (restriction), hsdM (modification) and hsdS (specificity subunit that binds to an asymmetrical Deoxyribonucleic acid sequence and determines the specificity of restriction and methylation). Two hsdM subunits and 1 hsdS subunit are involved in methylation of self-DNA. On the other paw, 2 complexes of hsdR, hsdM and hsdS (where each complex consists of two hsdR, two hsdM and one hsdS subunit) bind to the unmethylated recognition sites on phage DNA and cleave the DNA at random, far from their recognition sequences. Both reactions—methylation and cleavage—require ATP. (b) Type II R-M system is composed of two singled-out enzymes: palindromic sequence methylating methyltransferase (mod) and endonuclease (res) that cleave unmethylated palindromic sequences close to or inside the recognition sequence. (c) Type Iii R-Grand system is formed of methyltransferase (mod) and endonuclease (res) that course a complex. Methyltransferase transfers methyl group to one strand on the Dna, whereas 2 methyltransferases (endonuclease complexes) act together to bind to the complementary unmethylated recognition sites to cleave the Deoxyribonucleic acid 24–26 bp abroad from the recognition site. (d) Type IV R-Thou organization contains but endonuclease (res) that recognises methylated or modified Deoxyribonucleic acid. Cleavage occurs within or away from the recognition sequences [
Due to the diversity of R-M systems, phages acquired several agile and passive strategies to bypass cleavage by REases. Passive mechanisms include reduction in restriction sites, modification and change of the orientation of restriction sites, whereas more than specific, agile mechanisms include masking of restriction sites, stimulation of MTase activeness on phage genome or degradation of an R-Chiliad system cofactor (Figure 7) [fifteen, 16, 17, 18].

Figure vii.
Phage'due south passive and active strategies to bypass restriction-modification (R-M) systems. (a) Phages that possess fewer restriction sites in their genome are less prone to DNA cleavage by the host restriction endonuclease (REase). (b) Occasionally phage Dna might be modified by bacterial methyltransferase (MTase) upon successful injection into a host cell. Methylated recognition sites on viral DNA are, therefore, being protected from the cleavage and degradation past REase, leading to the initiation of the phage's lytic cycle. In addition, some phages encode their own MTase that is cooperative with the host REase; thus viral DNA cannot be recognised equally nonself. (c) Some phages, for case, coliphage P1, while injecting its DNA into a host cell, it also co-injects host-genome-binding proteins (DarA and DarB) that mask R-Thousand recognition sites. (d) Phages such equally Coliphage T7 possess proteins that can mimic the DNA courage. Ocr, a poly peptide expressed by Coliphage T7, mimics the DNA phosphate backbone and has a loftier affinity for the EcoKI REase component, thereby interfering with R-M system. (e) In addition, some phages (east.grand. Ral protein of Coliphage λ) can as well stimulate activity of the bacterial modification enzyme in lodge to protect own Dna from the recognition by the bacterial REase as nonself. The peptide Stp encoded past Coliphage T4 tin can besides disrupt the structural conformation of the REase-MTase complex [
Fewer brake sites in the evading genome lead to the selective advantage of this phage equally its DNA is less prone to cleavage and degradation by the host REase (Figure 7a). Also, some phages incorporate modified bases in their genomes that may lead to successful infection of the host prison cell as REase may not recognise the new sequences in the restriction sites. A decrease in the effective number of palindromic sites in DNA or change in the orientation of restriction-recognition sites can affect R-Thou targeting. Alternatively, the recognition sites within the viral genome can be too distant from each other to be recognised and cleaved past the REase [15, sixteen, 17, 18, 53].
Interestingly, phage genome might be methylated past bacterial MTase upon successful injection into a host cell. Methylated recognition sites on viral genomes are therefore being protected from the cleavage and degradation by REase, leading to the initiation of the phage's lytic cycle. Viral progeny remains insensitive to this specific bacterial REase until it infects a bacterium that possesses a different type of REase, in which instance the new progeny will become unmethylated again and will, therefore, be sensitive to the R-M arrangement of the cognate bacterium [28, 29, 30, 31].
The fate of the host cell chiefly confides in the levels of R-K gene expression and ultimate proportion of the R-M enzymes and their competition for the sites in the invading phage genome [52].
Furthermore, some phages encode their own MTase that is cooperative with the host REase, and thereby viral DNA cannot be recognised every bit nonself. Phages can as well stimulate the activeness of host modification enzymes that tin rapidly methylate viral Dna, thus protecting it from the activity of REase.
Alternatively, phages can bypass R-Chiliad systems by masking restriction sites. For example (Effigy 7c), coliphage P1, while injecting its Dna into a host prison cell, it also co-injects host-genome-binding proteins (DarA and DarB) that mask R-M recognition sites [53, 54].
As shown on an example of a Coliphage T7 (Figure 7d), some phages lawmaking for proteins that directly inhibit REase. Coliphage T7 possesses proteins that can mimic the Deoxyribonucleic acid courage. Ocr, a protein expressed past Coliphage T7, directly blocks the active site of some REases by mimicking 24 bp of bent B-form DNA, and it has a high affinity for the EcoKI REase component, thereby interfering with R-Yard arrangement [53].
Lastly, phage-bacteria arm race immune phages to proceeds capabilities of degrading necessary cofactors of R-Grand systems. For instance, coliphage T3 encodes S-adenosyl-50-methionine hydrolase that destroys an essential host R-Chiliad cofactor (the Southward-adenosyl-50-methionine). The removal of this necessary co-cistron will lead to the inhibition of the REase, thereby successfully infecting the host cell [15, xvi, 17, 18].
v.ii CRISPR/Cas arrangement
CRISPR forth with CRISPR-associated (Cas) proteins is the blazon of adaptive heritable 'immunity' of leaner, thus very specific and effective; and it is prevalent inside the bacterial domain [55]. The CRISPR are Dna loci consisting of curt palindromic repeats (identical in length and sequence), interspaced by segments of DNA sequences (spacer Dna) derived from previous exposures to phages. The spacer DNA sequences human activity as a 'retention', allowing bacteria to recognise and destroy specific phages in a subsequent infection. Genes encoding Cas proteins are adjacent to CRISPR loci [56].
Although some studies have suggested that CRISPRs can be used for pathogen subtyping [57], information technology has been found that CRISPR typing is not useful for the epidemiological surveillance and outbreak investigation of
The CRISPR/Cas phage resistance is mediated in 3-step stages: adaptation (acquisition), where spacer phage-derived DNA sequences are incorporated into the CRISPR/Cas system; expression, where
CRISPR/Cas systems have been classified into three major types: Types I, II and III, which are further divided into subtypes that crave different types of Cas proteins. Although the CRISPR/Cas array is various amidst the bacteria and information technology is continuously co-evolving in response to the host-phage interactions, the defence activity in all three types of the CRISPR is comparable [21, 22, 23] Figure 8 illustrates the defence mechanisms in iii distinct CRISPR/Cas arrays.

Effigy 8.
Prototype showing mechanisms of adaptation, expression and interference in iii different types of CRISPR/Cas arrays. Type I and Type II CRISPR/Cas arrays rely on the protospacer side by side motif (PAM), contained inside phage nucleic acid, to 'select' the phage-derived protospacer. Next steps in the adaptation phase are similar in all 3 types; protospacer is incorporated by Cas i and Cas2 proteins into the bacterial genome at the leader end of the CRISPR loci to grade a new spacer. In expression step, CRISPR loci are transcribed into pre-crRNA. The crRNA processing and interference stage is singled-out in each type of the CRISPR/Cas system. In Type I, the multisubunit CRISPR-associated circuitous for antiviral defence (Pour) binds crRNA to locate the target, and with the presence of Cas3 protein, the invading target genome is degraded whereas in Type II, Cas9 poly peptide is essential in the processing of the crRNA. TracrRNA recognises and attaches to the complementary sequences on the echo region that is and then cut by RNase III in the presence of Cas9. Lastly, in Type Three, processing of pre-crRNA into crRNA is dependent upon the activity of Cas6. Mature crRNA associated with Csm/Cmr complex targets foreign DNA or RNA for the degradation [
The Type II, CRISPR/Cas9, which was start identified in
In the adaptation phase, phage-derived protospacer (snippet of DNA from the invading phage) is incorporated into the bacterial genome at the leader end of the CRISPR loci. In expression phase, the
To bypass CRISPR/Cas that has an incredibly dynamic rate of evolution, phages caused assortment of strategies to succeed in propagation; this includes mutations in the protospacers or in the PAM sequences and expression of anti-CRISPR proteins, and fifty-fifty some phages encode their own functional CRISPR/Cas systems [15, 16, 17, 18, 63].
Phages can evade interference step of Type I and Type Two CRISPR/Cas system by a unmarried point mutation or deletion in their protospacer region or in the PAM sequence (Figure 9). Phages with single-nucleotide substitutions or deletions positioned close to PAM sequence can featherbed the CRISPR/Cas activity and consummate their lytic cycles; in contrast, phages with multiple mutations at PAM-distal protospacer positions do not [fifteen, 16, 17, 18, 28, 29, 30, 31].

Figure nine.
Evasion by mutation. Mutations in the phage protospacers or in the PAM sequences allow the phage to escape interference step of the CRISPR/Cas arrangement that would lead to the deposition of the phage genome [
In some circumstances, yet, although the phage successfully evades CRISPR/Cas interference, the host cell may survive by the acquisition of new spacer sequences (derived from invading phage) into their own CRISPR/Cas system. This new spacer provides the bacterium with an accelerated spectrum of phage resistance [xv, sixteen, 17, 18].
Prophages integrated within

Effigy ten.
Anti-CRISPR proteins expressed against CRISPR subtype I-F systems. Temperate phages such equally
Prophages practice not only contribute to bacterial resistance to invading phages, they tin can also encode proteins that contribute to bacterial virulence and antimicrobial resistance [58, 66].
Bacteria can besides resist phages by possessing phage-inducible chromosomal islands (PICI) which forestall phage replication. Nevertheless, phages evolved their genomes to overcome this very specific antiviral strategy. For instance,

Figure 11.
Phage-encoded CRISPR/Cas systems in
Advertizement
six. Overcoming host abortive infection systems: toxin-antitoxin
Abortive infection (Abi) systems promote prison cell decease of the phage-infected bacteria, inhibiting phage replication and providing protection for bacterial populations [68].
Abi systems require both toxins and antagonistic antitoxins. Antitoxins are proteins or RNAs that protect bacterial cell from the activity of toxins in a typical cell life cycle, whereas toxins are the proteins encoded in toxin-antitoxin locus that disrupt cellular metabolism (translation, replication and prison cell wall formation), causing cell death. During an infection, the expression of the antitoxin encoding cistron is suppressed, leading to the lethal activation of the toxin [69]. Figure 12 illustrates the mechanism of Abi systems in

Figure 12.
Bootless infection (Abi) systems in
Interestingly, phages evolved an array of tactics to circumvent Abi systems. This includes mutations in specific phage genes and encoding own antitoxin molecules that suppresses bacterial toxin [xv, sixteen, 17, 18]. Figure 13 provides a wide overview of the strategies employed by the phages to by-pass Abi systems.
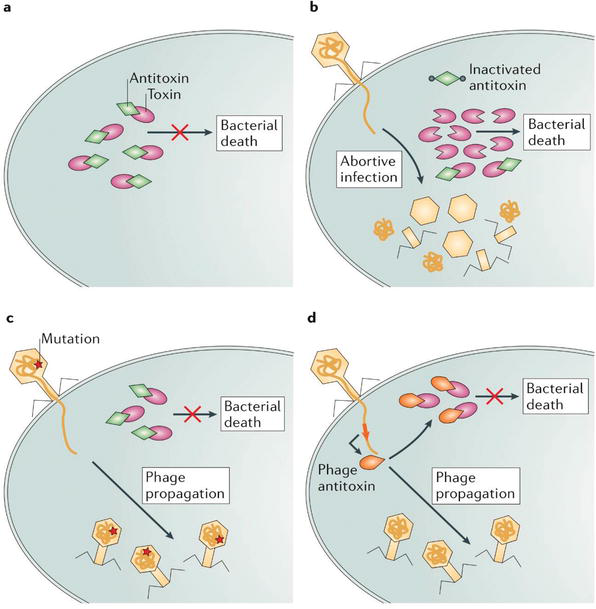
Figure thirteen.
Escaping abortive infection mechanisms. (a) In a typical cell life cycle, antitoxins protect bacterial cell from the activity of toxins. (b) During phage infection, the expression of antidote encoding cistron is suppressed, leading to the lethal activation of the toxin. (c) Mutations in sure phage genes tin can pb to escaping Abi systems activeness, thereby a successful viral propagation without killing the host cell. (d) Some phages encode molecules that functionally supersede the bacterial antitoxins, thus suppressing toxin activity and avoiding host cell death [
Bacteria-phage interaction is therefore very complex, and it is crucial to empathise the molecular basis of this interaction and how leaner and phages 'fight' each other. It has been reported that Anderson Phage Typing Arrangement of
Advertisement
7. The potential application of phages equally antibacterial therapeutics
The rapid emergence and dissemination of MDR bacteria seriously threaten global public health, as, without effective antibiotics, prevention and treatment of both community- and infirmary-caused infections may become unsuccessful and lead to widespread outbreaks.
Carbapenems and colistin are antibiotics of last resort, generally reserved to treat bacteria which are resistant to all other antibiotics. Until non long ago, colistin resistance was only described as chromosomal, nevertheless, in 2016 Liu et al. reported the emergence of the commencement plasmid-mediated colistin resistance mechanism, MCR-i, in Enterobacteriaceae [72]. Furthermore, the increasing occurrence of colistin resistance among carbapenem-resistant Enterobacteriaceae has also been reported [73]. This is of significant concern as infections caused by colistin and carbapenem-resistant leaner are very challenging to treat and command, every bit the treatment options are greatly limited or non-real. Thus, the discovery and development of alternative antimicrobial therapeutics are the highest priorities of modern medicine and biotechnology.
Phages should be considered as great potential tools in MDR pathogens as they are species-specific (specificity prevents damage of normal microbiota), thus harmless to homo; they take fast replication rate at the site of infection, and their short genomes can allow to further understand various molecular mechanisms implied to 'fight' bacteria. In improver, this understanding can enable scientists to 'manipulate' viral genomes and engineer a synthetic phage that combines the antibacterial characteristics of multiple phages into a single genome.
The escalating need for new antimicrobial agents attracted new attention in modern medicine, proposing several potential applications of phages as antibacterial therapeutics including phage therapy, phage lysins and genetically-engineered phages.
7.1 Phage therapy
Phage therapy utilises strictly lytic phages that have bactericidal effect. Equally phages are host-specific, 'phage cocktails' containing multiple phages tin augment range of target cells. Withal, selection of suitable phages is at the paramount to the successful elimination of clinically of import pathogens, and it includes avoidance of adverse effects, such equally anaphylaxis (adverse immune reaction) [74].
7.2 Phage-derived enzymes: lysins
In order to hydrolyse and degrade the bacterial cell wall, phages possess lysins.
The spectrum of efficiency of natural lysins (derived from naturally occurring phages) is generally limited to Gram-positive leaner; still, recombinant lysins have shown an ability to destabilise the outer membrane of Gram-negative bacteria and ultimately lead to rapid death of the target bacteria [74].
7.3 Bioengineered phages
Bioengineered phages have the potential to solve inherent limitations of natural phages such every bit narrow host range and development of resistance. Various genetic applied science methods accept been proposed to design phages with extended antimicrobial properties such as homologous recombination, phage recombineering of electroporated Dna, yeast-based platform, Gibson assembly and CRISPR/Cas genome editing [75].
Applied science of constructed phages could be tailored to raise the antibiotic activeness, to reverse antibiotic resistance or to create sequence-specific antimicrobials [74].
Advertisement
8. Conclusions
The combative host-phage relationship has led to the evolution of uncommonly disperse phage-resistance mechanisms in the bacterial domain, including inhibition of phage adsorption, prevention of nucleic acrid entry, Superinfection exclusion, cutting phage nucleic acids via restriction-modification systems and CRISPR, as well as bootless infection.
Evolvement of these mechanisms has been induced by abiding parallel co-evolution of phages as they effort to coexist. To survive, phages acquired diverse counterstrategies to circumvent bacterial anti-phage mechanisms such as adaptations to new receptors, digging for receptors and masking and modification of restriction sites and point mutations in specific genes and genome rearrangements that allow phages to evade bacterial antiviral systems such as CRISPR/Cas arrays, besides every bit mutations in specific genes to bypass abortive infection arrangement. Conclusively, the co-evolving genetic variations and counteradaptations, in both leaner and phages, drive the evolutionary bacteria-host arm race.
Besides, accumulating testify shows that phages contribute to the antimicrobial resistance through horizontal gene transfer mechanisms. Indeed, many bacterial strains have become insensitive to the conventional antibiotics, posing a growing threat to human; and although in the past, western counties withdrew phage therapy in response to the discovery of therapeutic antibiotics, now, phage therapy regains an involvement within the research customs. In that location are apparent advantages of phage therapy, such equally specificity, pregnant only target leaner would encounter lysis, only not salubrious microbiota inhabiting human's system. Additionally, 'phage cocktails', containing multiple leaner-specific phages, could overcome the issue of phage-resistance equally phages do suit to these resistance mechanisms. However, 'phage cocktails' would require large numbers of phages that would have to exist grown inside pathogenic leaner in the laboratory, putting laboratory staff and the surround at risk.
Alternatively, building up the understanding of host-phage interactions and 'the war between leaner and phages' could potentially lead to defeating antimicrobial resistance by designing constructed phages that can overcome the limitations of phage therapy.
Advertisement
Acknowledgments
Dr Manal Mohammed is funded by a Quinton Hogg starting time-up award, University of Westminster.
Advertisement
Abbreviations
abortive infection
capsular polysaccharides
clustered regularly interspaced brusque palindromic repeats
crispr RNA
diversity-generating retroelement
deoxyribonucleic acid
multidrug-resistant
O-methyl phosphoramidate
methyltransferase
protospacer adjacent motif
phage-inducible chromosomal island
PICI-like chemical element
receptor-bounden protein
restriction endonuclease
restriction-modification
ribonucleic acid
superinfection exclusion
trans-activating crRNA
References
- 1.
Public Health England. Health Matters: Antimicrobial Resistance. Government Digital Service. 2018. Bachelor from: https://world wide web.gov.uk/authorities/publications/wellness-matters-antimicrobial-resistance/health-matters-antimicrobial-resistance [Accessed: 10 July 2018] - ii.
Slama TG. Gram-negative antibiotic resistance: There is a toll to pay. Disquisitional Care. 2008; 12 :S4 - 3.
Herelle FD. The Bacteriophage: Its Rôle in Immunity. English ed. Baltimore: Williams & Wilkins; 1922 - 4.
Sulakvelidze A, Alavidze Z, Morris JG. Bacteriophage therapy. Antimicrobial Agents and Chemotherapy. 2001; 45 (3):649 - 5.
Fauconnier A. Regulating phage therapy. EMBO Reports. 2017; 18 (2):198-200 - six.
Baharuddin A et al. Revitalizing phage therapy in combating multi-drug resistant bacteria. Haya: The Saudi Journal of Life Sciences. 2017; ii (4):122-130 - 7.
Bikard D, Marraffini LA. Innate and adaptive immunity in bacteria: Mechanisms of programmed genetic variation to fight bacteriophages. Current Opinion in Immunology. 2012; 24 (1):xv-twenty - viii.
Breitbart M, Rohwer F. Here a virus, there a virus, everywhere the same virus? Trends in Microbiology. 2005; thirteen (vi):278-284 - nine.
Muhammad Abu BS et al. Leaner vs. bacteriophages: Parallel development of immune arsenals. Frontiers in Microbiology. 2016; seven :1292 - ten.
Chao L, Levin BR, Stewart FM. A complex community in a simple habitat: An experimental study with bacteria and phage. Ecology. 1977; 58 (2):369-378 - xi.
Bohannan B, Lenski R. Linking genetic alter to community evolution: Insights from studies of bacteria and bacteriophage. Environmental Messages. 2000; 3 (4):362-377 - 12.
Chibani-Chennoufi South, Bruttin A, Dillmann M, Brussow H. Phage-host interaction: An ecological perspective. The Journal of Bacteriology. 2004; 186 (12):3677 - xiii.
Van Valen L. A new evolutionary constabulary. Evolutionary Theory. 1973; ane (ane):i-thirty - 14.
Betts A et al. Assorted coevolutionary dynamics between a bacterial pathogen and its bacteriophages. Proceedings of the National Academy of Sciences. 2014; 111 (30):11109 - fifteen.
Samson JE et al. Passive and agile strategies to avert restriction-modification systems [image]. 2013. Bachelor from: https://www.nature.com/articles/nrmicro3096 [Accessed: 9 July 2018] - 16.
Samson JE et al. Phage strategies to by-pass CRISPR-Cas systems [image]. 2013. Available from: https://www.nature.com/articles/nrmicro3096 [Accessed: ix July 2018] - 17.
Samson JE et al. Phage strategies to past-pass toxin–antitoxin systems [image]. 2013. Bachelor from: https://world wide web.nature.com/articles/nrmicro3096 [Accessed: eleven July 2018] - 18.
Samson JE et al. Revenge of the phages: Defeating bacterial defences. Nature Reviews Microbiology. 2013; xi (10):675 - 19.
Hyman P, Abedon ST. Bacteriophage host range and bacterial resistance—chapter seven. Advances in Applied Microbiology. 2010; 70 :217-248 - 20.
Doron S et al. Systematic discovery of antiphage defense systems in the microbial pangenome. Science (New York, N.Y.). 2018; 359 (6379):eaar4120 - 21.
Makarova KS et al. Development and classification of the CRISPR-Cas systems. Nature Reviews. Microbiology. 2011; ix (half-dozen):467 - 22.
Makarova KS et al. Defence force Islands in bacterial and Archaeal genomes and prediction of novel defense systems. Journal of Bacteriology. 2011; 193 (21):6039-6056 - 23.
Makarova KS et al. The three stages of CRISPR–Cas action [paradigm]. 2011. Available from: https://www.nature.com/manufactures/nrmicro2577 [Accessed: eight July 2018] - 24.
Vale PF, Little TJ. CRISPR-mediated phage resistance and the ghost of coevolution by. Proceedings of the Royal Club B. 2010; 277 (1691):2097-2103 - 25.
Etz H et al. Bacterial phage receptors, versatile tools for display of polypeptides on the prison cell surface. The Journal of Bacteriology. 2001; 183 (23):6924 - 26.
Dy RL et al. The mechanisms for methylation and restriction for the 4 types of brake-modification (RM) systems [epitome]. 2014. Bachelor from: https://www.annualreviews.org/doi/abs/10.1146/annurev-virology-031413-085500?journalCode=virology [Accessed: 8 July 2018] - 27.
Dy RL et al. Remarkable mechanisms in microbes to resist phage infections. Annual Review of Virology. 2014; 1 :307-331 - 28.
Labrie SJ, Samson JE, Moineau S. Bacteriophage resistance mechanisms. Nature Reviews Microbiology. 2010; viii (5):317 - 29.
Labrie SJ et al. Blocking phage DNA entry into the bacterial cell [image]. 2010. Bachelor from: https://www.ncbi.nlm.nih.gov/pubmed/20348932 [Accessed: 9 July 2018] - thirty.
Labrie SJ et al. Different strategies used by bacteria to cake phage adsorption [image]. 2010. Available from: https://world wide web.ncbi.nlm.nih.gov/pubmed/20348932 [Accessed: ix July 2018] - 31.
Labrie SJ et al. The fashion of activeness of the king system [prototype]. 2010. Bachelor from: https://www.ncbi.nlm.nih.gov/pubmed/20348932 [Accessed: vi July 2018] - 32.
Golais F, Hollý J, Vítkovská J. Coevolution of bacteria and their viruses. Folia Microbiologica. 2013; 58 (3):177-186 - 33.
Ahmad S et al. An overview on stage variation, mechanisms and roles in bacterial accommodation. JPMA. The Journal of the Pakistan Medical Association. 2017; 67 (2):285-291 - 34.
Guo H et al. Diversity-generating retroelements in phage and bacterial genomes. Microbiology Spectrum. 2014; 2 (6):2014 - 35.
Sørensen MC et al. The F336 bacteriophage recognizes the capsular phosphoramidate modification of Campylobacter jejuni NCTC11168. Periodical of Bacteriology. 2011; 193 :6742-6749. DOI: 10.1128/JB.05276-11 - 36.
Sørensen MC et al. Phase variable expression of capsular polysaccharide modifications allows Campylobacter jejuni to avoid bacteriophage infection in chickens. Frontiers in Cellular and Infection Microbiology. 2012; ii :11. DOI: ten.3389/fcimb.2012.00011 - 37.
Dai Due west et al. Iii-dimensional construction of tropism-switching Bordetella bacteriophage. Proceedings of the National University of Sciences of the United States of America. 2010; 107 (9):4347 - 38.
Nordstrom K, Forsgren A. Effect of protein a on adsorption of bacteriophages to Staphylococcus aureus . The Journal of Virology. 1974;14 (2):198 - 39.
Nwodo U, Light-green E, Okoh A. Bacterial Exopolysaccharides: Functionality and Prospects. Basel: MDPI AG; 2012 - 40.
Looijesteijn PJ et al. Physiological office of exopolysaccharides produced by Lactococcus lactis . International Journal of Food Microbiology. 2001;64 (1):71-lxxx - 41.
Sutherland IW. Polysaccharide lyases. FEMS Microbiology Reviews. 1995; sixteen (4):323 - 42.
Letarov A, Kulikov Due east. Adsorption of bacteriophages on bacterial cells. Biochemistry (Moscow). 2017; 82 (thirteen):1632-1658 - 43.
Orlova EV. How viruses infect bacteria? EMBO Journal. 2009; 28 (7):797-798 - 44.
Campbell A. Life wheel of the typical temperate phage coliphage-λ [image]. 2003. Available from: https://world wide web.nature.com/articles/nrg1089 [Accessed: vii July 2018] - 45.
Campbell A. The future of bacteriophage biology. Nature Reviews Genetics. 2003; 4 (6):471 - 46.
St-Pierrea F, Endy D. Conclusion of cell fate option during phage lambda infection. Proceedings of the National Academy of Sciences of the Us of America. 2008; 105 (52):20705-20710 - 47.
Folimonova SY. Superinfection exclusion is an active virus-controlled function that requires a specific viral poly peptide. Journal of Virology. 2012; 86 (10):5554 - 48.
Hofer B, Ruge M, Dreiseikelmann B. The superinfection exclusion gene (sieA) of bacteriophage P22: Identification and overexpression of the gene and localization of the gene product. The Journal of Bacteriology. 1995; 177 (11):3080 - 49.
Mohammed M, Cormican M. Whole genome sequencing provides possible explanations for the deviation in phage susceptibility amongst 2 Salmonella Typhimurium phage types (DT8 and DT30) associated with a unmarried foodborne outbreak. BMC Research Notes. 2015; 8 (1):728 - 50.
Vasu K, Nagaraja Five. Various Functions of Restriction-Modification Systems in Add-on to Cellular Defense force. Washington: American Guild for Microbiology; 2013 - 51.
Vasu 1000, Nagaraja V. Brake-modification (R-M) systems as defence mechanisms [image]. 2013. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3591985/ [Accessed: 5 July 2018] - 52.
Enikeeva FN et al. Restriction-modification systems and bacteriophage invasion: Who wins? Journal of Theoretical Biology. 2010; 266 (iv):550-559 - 53.
Stern A, Sorek R. The phage-host arms race: Shaping the evolution of microbes. BioEssays. 2011; 33 (1):43-51 - 54.
Tock MR, Dryden DT. The biological science of restriction and anti-brake. Electric current Opinion in Microbiology. 2005; 8 (4):466-472 - 55.
Marchfelder A. Special focus CRISPR-Cas. RNA Biology. 2013; 10 (5):655-658 - 56.
Heler R et al. Cas9 specifies functional viral targets during CRISPR-Cas adaptation. Nature. 2015; 519 (7542):202L - 57.
Fabre Fifty et al. CRISPR typing and subtyping for improved laboratory surveillance of salmonella infections (CRISPR polymorphisms in salmonella). PLoS I. 2012; 7 (5):e36995 - 58.
Mohammed Thousand et al. The invasome of Salmonella Dublin as revealed past whole genome sequencing. BMC Infectious Diseases. 2017; 17 (1):544 - 59.
Koonin EV, Makarova KS. CRISPR-Cas: Evolution of an RNA-based adaptive immunity arrangement in prokaryotes. RNA Biology. 2013, 2013; 10 (5):679-686 - 60.
Leon LM, Mendoza SD, Bondy-Denomy J. How bacteria command the CRISPR-Cas arsenal. Current Opinion in Microbiology. 2018; 42 :87-95 - 61.
Ka D et al. Crystal structure of streptococcus pyogenes Cas1 and its interaction with Csn2 in the type 2 CRISPR-Cas system. Structure. 2016; 24 (1):lxx-79 - 62.
Carroll D. Staying on target with CRISPR-Cas. Nature Biotechnology. 2013; 31 (ix):807-809 - 63.
Mohanraju P et al. Diverse evolutionary roots and mechanistic variations of the CRISPR-Cas systems. Science. 2016; 353 (6299) - 64.
Wiedenheft B. In defense of phage: Viral suppressors of CRISPR-mediated adaptive immunity in bacteria. RNA Biology. 2013; 10 (5):886 - 65.
Seed K et al. A bacteriophage encodes its own CRISPR/Cas adaptive response to evade host innate amnesty. Nature. 2013; 494 (7438):489-491 - 66.
Mohammed M, Cormican M. Whole genome sequencing provides insights into the genetic determinants of invasiveness in Salmonella Dublin. Epidemiology and Infection. 2016; 144 (11):2430-2439 - 67.
Naser I et al. Assay of the CRISPR-Cas system in bacteriophages active on epidemic strains of Vibrio cholerae in People's republic of bangladesh. Scientific Reports. 2017;vii (1):14880 - 68.
Chopin 1000, Chopin A, Bidnenko E. Phage abortive infection in lactococci: Variations on a theme. Current Opinion in Microbiology. 2005; viii (4):473-479 - 69.
Fineran PC et al. The phage bootless infection organisation, ToxIN, functions equally a poly peptide-RNA toxin-antitoxin pair. Proceedings of the National Academy of Sciences of the United states of america. 2009; 106 (3):894 - 70.
Refardt D, Kümmerli R. Defying bacteriophages: Contrasting donating with individual-based resistance mechanisms in Escherichia coli . Communicative & Integrative Biology. 2013;6 (5):e25159 - 71.
Mohammed M. Who fights whom? Understanding the complex dynamics of bacteria-phage interaction using Anderson phage typing system. Journal of Infectious Diseases & Therapy. 2018; vi (iii):367 - 72.
Liu Y et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and man beings in China: A microbiological and molecular biological study. The Lancet Infectious Diseases. 2016; sixteen (two):161-168 - 73.
European Eye for Illness Prevention and Command. Expert consensus protocol on colistin resistance detection and characterisation for the survey of carbapenem- and/or colistin-resistant Enterobacteriaceae. 2019. Available from: https://ecdc.europa.eu/en/publications-data/expert-consensus-protocol-colistin-resistance-detection-and-characterisation [Accessed: thirteen May 2019] - 74.
Kakasis A, Panitsa G. Bacteriophage therapy as an alternative treatment for human infections. A comprehensive review. International Journal of Antimicrobial Agents. 2019; 53 (i):16-21 - 75.
Monteiro R et al. Phage therapy: Going temperate? Trends in Microbiology. 2019; 27 (four):368-378. DOI: x.1016/j.tim.2018.10.008
Submitted: July 15th, 2018 Reviewed: June 4th, 2019 Published: October 31st, 2019
© 2019 The Author(south). Licensee IntechOpen. This chapter is distributed under the terms of the Creative Eatables Attribution iii.0 License, which permits unrestricted use, distribution, and reproduction in whatever medium, provided the original piece of work is properly cited.
Source: https://www.intechopen.com/chapters/69255
0 Response to "Bacterial Defence Mechanisms and Bacteriophage Counter-defence Mechanisms Nature Review Microbiology"
Post a Comment